显微镜对RNA的研究
什么是RNA?
核糖核酸简称RNA。 这些分子是必不可少的生活的几乎所有的工序,因为它们介导的所有步骤的基因表达的:信使RNA(mRNA)从基因转录,携带信息出细胞核。 在转录真核生物中,mRNA的成熟,其中涉及拆除插序列(内含子)的。 这个过程 - 被称为拼接 - 主要是通过SN /含üRNA剪接体介导的。 在从核出口,大部分的mRNA翻译得到的含rRNA的核糖体 - 该代码是通过携带激活的氨基酸合成蛋白质转运RNA解释。 微RNA,短干扰RNA和的piRNA - 翻译和特定mRNA的寿命部分由短RNA控制的。 不仅上述的RNA种类,但实际上该基因组的大部分被转录 - 其中的意义目前正在积极研究。
什么是显微镜都与RNA?
近三十年前,被发现β-actin基因分子被不均匀鸡成肌细胞和成纤维细胞的细胞质中本地化。 很快,在其他模型系统 - 许多其他的例子,如芽殖酵母,果蝇和非洲爪蟾卵母细胞和哺乳动物成纤维细胞和神经元 - 紧随其后。 Zui近,在果蝇胚胎和卵母细胞的RNA本地化的全基因组的研究表明,所表达的转录物的大部分(高达80%)分布于细胞质内的独特的,非均匀的图案。 此外,尽管是微小的,没有明确定义的细胞器,细菌被证实为表达本地化的表达。
将近十年前,所推出的单分子敏感性显微技术,它成为可以测量绝对RNA的数量 - 通过计算RNA分子 - 以前所未有的细胞或亚分辨率。 这些技术是目前允许各个单元的“深转录组测序”的显微镜载物台的边缘。
当然,显微镜,不仅提供了快照 - 位置和号码 - RNA分子,但有合适的成像方法,转录深刻的机制,RNA本地化,翻译和衰变被发现。
你怎么能想象RNA?
核糖核酸 - 类似于DNA和多肽 - 是聚合物大分子。 其身份是由主结构或积木序列中,核糖核苷酸(A,C,G和U)来决定。 大多数的RNA分子是单链的 - 虽然它们折叠成复杂的二级和三级结构往往涉及形成分子内双链体 - 从而他们的身份可以与匹配互补序列进行探测。 这些探头大多是单链自然的核酸 - 单链DNA或RNA - 携带一些检测标记,从放射性同位素,小,免疫检测的分子 - haptenes - 或荧光染料。 期间称为原位杂交(ISH)的方法,该探针找到并结合其靶RNA。 不匹配的无杂交通常更稳定能量上比从而通过在一个特定信号被开发的ISH方法适当的洗涤步骤被探针形成的,其他的,非特异性复合物是。 然而,由于这些强制性分化步骤: - 在恶劣治疗旁一些ISH协议要求(使用高温,有机溶剂,高盐等) - 没有这些ISH技术是与生活细胞成像相容。
那你怎么跟着RNA住在哪里?
RNA永远存在“裸”细胞内的 - 它总是在复杂的一组动态的蛋白质分子形成核糖核蛋白颗粒(的RNP)。 许多这些蛋白质分子的结合的核糖核酸直接(限制性商业惯例)。 融合这些限制性商业惯例到荧光蛋白(FPS)是一种广泛使用的方法来可视化的RNA活。 大多数这些限制性商业惯例,在真核细胞的剧目发现 - 例如诗道芬,eIF4AIII,Hrp48A,PABP等,限制了目标特异性和任何给定的RNP只是兼职居民。 然而一些限制性商业惯例,如Pumillio同源域(PUM-HD)或CRISPR / Cas9,可以工程化以匹配给定的RNA靶。 然而,为了有效地显现mRNPs - 通常含有单个mRNA分子 - 每个粒子几个拷贝的RBP-FP融合体是必要的。 为了Zui大限度地减少每靶分子转基因构建体的数量,在体内的RNA可视化技术的Zui常用的利用限制性商业惯例直系到主机。 第一他们的种类是MS2系统,由6-24份的RNA茎环结构的特定于MS2噬菌体和匹配的MS2外壳蛋白融合至FP串联排列的。 通过引入MS2循环数组复制到目标ASH1 mRNA和封存未绑定的MCP-FP到细胞核,歌手实验室可以想像生活中的芽殖酵母动态ASH1 mRNPs。
这些噬菌体环/环结合外壳蛋白系统的一个共同的缺点是它们依赖于RNA靶的转基因修饰 - 大多引入另外的额外拷贝的内源性本两个等位基因 - 和回路装入外套阵列蛋白加入约1.5-2丙二醛到感兴趣的RNP的大小。 这些变化可能会影响目标限制性商业惯例的动力(转录,运输,翻译和衰减),因此,理想情况下,所得到的结果应该与其他独立,互补的方法进行验证。
然后,还必须有其他的方法来遵循的RNP生活。 他们如何从以前的有什么不同?
1996年,特亚吉实验室介绍分子信标(MBS),率先采用荧光探针。 MB都是短寡核苷酸探针用荧光团和淬灭剂对,在其两个相对的末端。 这些末端(通常为6-7个核苷酸)是彼此互补的形成发夹结构从而定位所述荧光团旁边的猝灭剂(暗状态)。 于MB的目标特定中间部分(环)的杂交,所述发夹的茎被熔化,分离荧光团和淬灭剂,从而产生活化的荧光(亮状态)。 使用的MB - 荧光探针在一般的 - 不要求区分洗涤步骤和目标的无转基因修饰。
不幸的是,RNA / DNA杂交也不双链RNA 在体内稳定。 为了解决这个问题,这些探头的骨干进行化学修饰以隐藏细胞内核酸形成的混合体。 作为正副作用,这些修改 - phosphoribose骨干大多2'O甲基化和/或锁核酸掺入 - 增加的混合动力车允许较短的探针的稳定性。
FIT探头
什么是FIT代表什么?
噻唑橙(FIT)的强制插是另一种方式来获得荧光探针。 这些探头,由Seitz的实验室开发,包括嵌入DNA染料,噻唑橙(TO),为碱替代 - 即更换非端子座中的寡核苷酸序列。 这种染料是一种低粘度的环境的非荧光时,例如在溶液(暗状态)。 在探针的杂交,两侧碱基对的氢键大大限制了染料的扭转运动,从而迫使它变成像状态的插层,活化荧光(亮状态)。 其结果是,FIT探针 - 不像的MB - 对不匹配非常敏感 - 特别是对那些在于:紧邻。
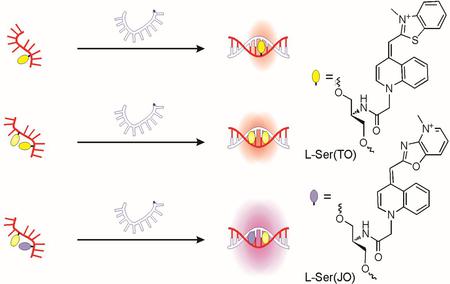
图1:(一)单一到,(B)增加一倍,和(C)东条标记的DNA FIT-探针的示意图。 资料来源:Hövelmann等人,2013。
做这些散客探头“样子”?
一个荧光报道的Zui重要的性质是亮度。 于:(量子产率)的发射在很大程度上取决于它的微环境; 不仅在探针的杂交的状态,而且还对位置和的邻居核苷。 在明亮的状态下,FIT探针杂交互补的单链DNA分子的典型亮度大约是7-14毫升⋅摩尔-1⋅-1,周围20-40%的的绿色荧光蛋白和AlexaFluor 488的10-20%。然而,成像的另一个关键参数是此相反,即信号到其直接背景的比值。 虽然传统的荧光团 - 像绿色荧光蛋白或AlexaFluor 488 - 都同样明亮独立地绑定到目标或解决方案,以黑暗状态标记FIT探针是8-20倍,比他们的明亮状态(响应)调光器。 此属性使FIT探针适于例如执行原位杂交(FISH),即使在其他方式挑战组织例如果蝇子房快速洗无荧光的。
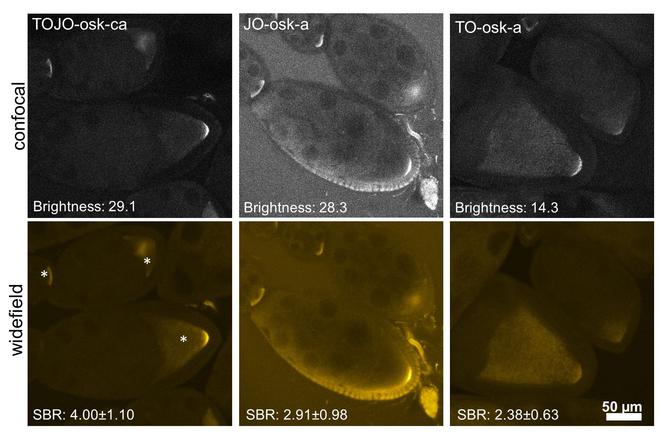
图2: 奥斯卡 mRNA在显影果蝇卵室清洗-自由FISH:蛋室包含显影卵母细胞(*),可以丰富奥斯卡的mRNAZui终在后极(在asterixes的右侧)。 该mRNA许多人一样是15护士细胞(左到卵母细胞)的乘积。 这16种系细胞被包封的体细胞滤泡epithelium.Source的单层:Hövelmann,2013。
您可通过将多个每个探头染料更亮FIT探头?
不是真的 - 因为这些探针是短,典型地15-30个核苷酸长(5-10纳米),两个或更多的荧光团被定位以及荧光共振能量转移的(FRET)的范围内,从而淬火彼此,导致只有20亮度增加-40%。 但是,我们设法增加一倍的亮度(约24-28毫升⋅摩尔-1⋅-1)通过引入一个稍微红移和明亮的荧光,恶唑(JO)进入到包含FIT探头。 JO,而被嵌入染料,是相当明亮,即使在溶液中; 这是它具有低响应速度。 在单链探针,然而,为了有效地猝灭JO碰撞。 杂交导致亮状态。 由于所形成的双链体的刚性,TO不再猝灭JO。 相反,激发能量通过FRET传递从TO到JO,恢复JO的Zui大亮度。
另一选项以增加单个FIT探针的亮度是增加所经历到本地约束。 我们表明,引入锁核酸(LNA)的3',以加倍的给定探针的Zui大亮度,同时它离开响应有效不变。 这种效应是由于到LNA引起的减少堆垛层相邻碱基之间的距离在双面增加当地粘度。
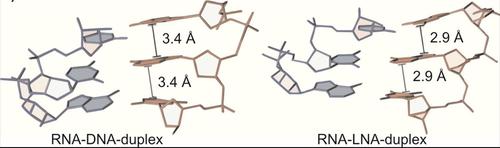
图3:由LNA减少堆叠距离增加本地粘度。 资料来源:Hövelmann和加斯帕,2014年。
你可以用这样的亮度提高了适配性探头的RNP体内成像?
几乎-在探头骨干单一LNA的修改并没有赋予足够的核酸酶抵抗形成复式然而,与核糖(mixmer设计)的2'OH额外的甲基化,探头被注入果蝇后保持稳定了一个多小时卵母细胞。 此外,一个探头很少是足够明亮,显示个别的RNP。 通过将三至五个不同的mixmer探头对奥斯卡的mRNA,我们设法获得亮度和对比度相似,10倍MS2标签标有MCP-EGFP转基因奥斯卡的RNP。
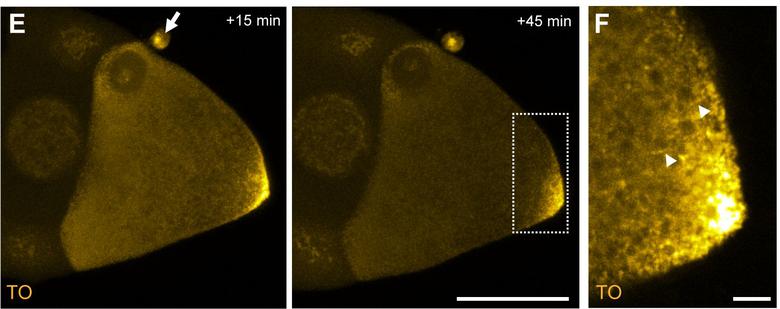
4A
图4:RNA体内成像:靶向奥斯卡 mRNA的3 mixmer FIT探头的混合物注入生活,野生型卵母细胞(G,箭头标记) 奥斯卡 RNP运动(H,箭头)可以遵循后甚至50分钟。注射。 ,比例尺代表50微米和5微米的G和H分别。 资料来源:Hövelmann和加斯帕,2014年。
你怎么知道注射的探头还没有目标上的影响?
我们使用上述oskarMS2(10X)/ MCP-EGFP系统,我们和其他人有据可查,作为参考。 同时注入所有五个mixmer探测即使,我们观察到奥斯卡 RNP活力无显着差异相比,oskarMS2。 当这样一个独立的分析是不可用时,一个很好的做法可以测试多种不同的FIT探头不同的组合。
当然,并不是所有的探头是惰性的。 通过靶向已知双链二级结构为奥斯卡 mRNA定位(一个所谓的定位元件)与mixmer探针互补的链(osk6)之一重要,我们诱导的奥斯卡 RNP蠕动相同的缺陷如先前的报导一系列突变奥斯卡转基因的。 所述基于FIT探针操纵不仅少得多耗时比产生和分析的RNA的转基因拷贝,而且只有在RNP生物合成的特定步骤的效果 - 在运输,在这种情况下。 突变,在另一方面,可能影响从转录命运mRNA的开始,并且因此,所观察到的表型可能是由于一系列的缺陷的总和。
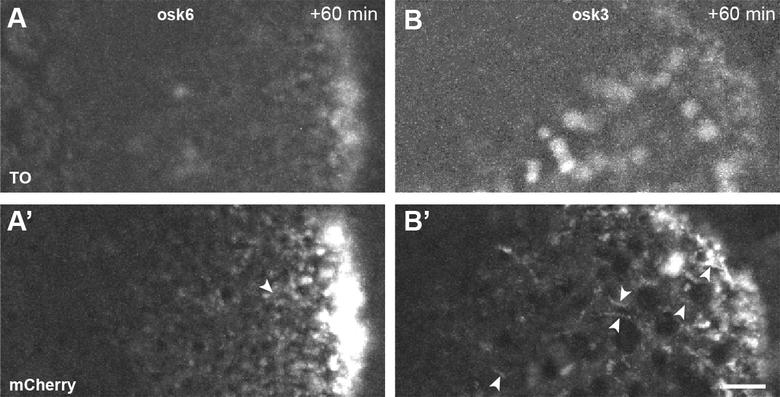
图5:干扰使用RNP功能FIT探头:本osk6 FIT探测目标的拼接奥斯卡本地化元素。 正如看到的TO通道(A和B)一个探头不够足够明亮按照个人的RNP,但是,它会报告成功定位。 上的时间突起oskarMS2(10倍)/ MCP-mCherry(A'和B')不存在长单向奥斯卡 mRNPs(箭头)的游程可以理解。 比例尺条为5微米。 资料来源:Hövelmann和加斯帕,2014年。
总之,仔细选择FIT探针是RNA可视化不仅是一种工具,但也RNP功能的发现。
您可以将这些探头与其他活细胞标记,如FP标签蛋白?
是的,可以。 由于大多数模式生物必须通过(E)绿色荧光蛋白标记的融合蛋白目前庞大的图书馆,这是至关重要的我们找到条件下,这两个荧光团 - EGFP和TO - 可以共同观察。 相比于:绿色荧光蛋白的吸收和发射光谱是足够红移,与仔细地选择激发光 - 470毫微米为EGFP和525纳米为TO - 有几乎在一个顺序扫描的两种分子之间没有串扰。 这些激发波长似乎有点异国风情 - 事实上,这样的双彩色成像,我们用一个国家的Zui先进的徕卡共聚焦显微镜配备了超连续光源。 自然,我们首先使用了oskarMS2(10倍)/ MCP-EGFP系统与FIT探针组合来检查用于共标记和两个信号的共迁移。
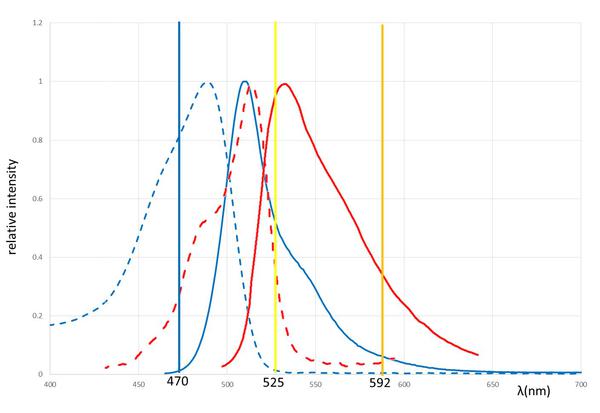
图6:吸收和GFP的发射光谱和TO:吸收(虚线)和发射(实线)的GFP(蓝色)和TO(红色)的光谱。 的激发和耗尽激光线波长由垂直实线表示。
更有趣的应用,但是,是测试的共定位用期间的RNP的生物合成,可能有功能的蛋白质。 经典,这些发现在连接质量生化分析完成。 镜,然而,有明显的优点超过这样的分析:它可以定义的地点,时间及有关的相互作用的动力学。 不幸的是,体RNP通常比衍射极限小,只包含少数 - 也许只有一个副本 - 一个给定的限制性商业惯例的。 因此,大多数免疫染色势必造成点状非特异性标记 - 即使特异性抗体 - 真正的RNP来自非特异性的背景难以区分。 EGFP融合蛋白的自发荧光,在另一方面,有一个几乎百分之百的特异性和少至单个GFP分子可与当今Zui灵敏的检测器来检测。
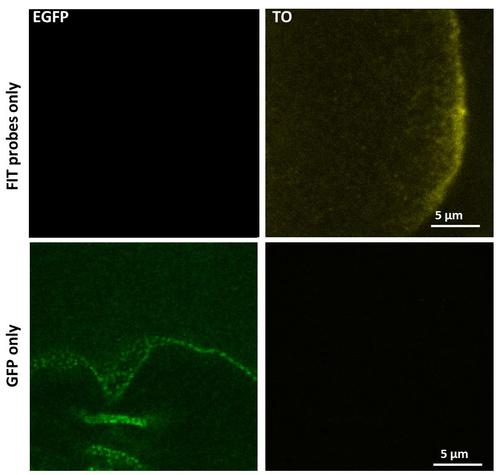
图7:瞄准奥斯卡 mRNA和核周GFP FIT探针成像单独使用相同的成像设置:GFP之间串扰。 有几乎EGFP和TO之间无串扰。 资料来源:伊姆雷·加斯帕,未公布。
如果体RNP是如此之小,你有没有考虑到的超高分辨率成像?
事实上 - 主要不是因为RNP大小,但因为拥挤,他们可以在某些位置实现。 因为果蝇卵室是一个相当厚的样品(100微米沿轴向轴)和有趣的部分是从玻璃表面通过10-15微米厚的层细胞的分离,这限制了方法的选择。我们发现,STED显微镜这些条件下工作。 LNA改性FIT探针已被证明是优良的受激发射损耗的标签,因为在杂交时不仅它们的亮度,而且荧光寿命显着增加,并且因此门控STED进一步提高了检测的信号的靶特异性。 因为在EGFP和TO的发射光谱重叠的同时,一个单个592纳米的激光耗竭足以实现双色STED成像。 虽然样本范围Zui大横向分辨率为约80-120纳米,无论是个人的RNP和RNP子光散射决心做更详细比传统的共聚焦显微镜。 这种成像使我们能够进行精确的基于对象的共定位分析。

图8:奥斯卡 mRNPs的STED成像:五LNA修饰FIT探针的混合物用于固定的果蝇卵巢。 中间oogenetic卵母细胞后极进行成像共聚焦,然后用门控STED设置。RAW图像进行反褶积与惠更斯专业。资料来源:伊姆雷·加斯帕,未公布。
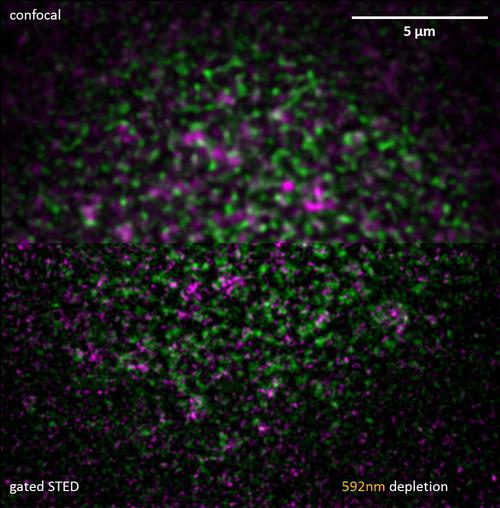
图9:双色STED:标有核周型绿色荧光蛋白(绿色)和瞄准奥斯卡的mRNA(品红色)FIT探头护士细胞的核信封。资料来源:伊姆雷·加斯帕,未公布。